

Мембраны биологические
МЕМБРАНЫ БИОЛОГИЧЕСКИЕ
(от лат. membrana-кожица, перепонка), сложные высокоорганизованные надмоле-кулярные
структуры, ограничивающие клетки (клеточные, или плазматич., мембраны) и внутриклеточные
органоиды -митохондрии, хлоропласты, лизосомы и др. Представляют собой пленки
толщиной 5-10 нм, состоящие гл. обр. из белков и липидов. Отношение липиды:
белки (по массе) колеблется от 4:1 (мембрана миелина) до 1:3 (внутр. мембрана
митохондрий). М. б. содержат также углеводы (до 10% от сухого в-ва по массе),
к-рые, как правило, входят в состав гликопротеинов и гликолипидов. В нек-рых
специали-зир. М.б. в заметных кол-вах могут присутствовать также хиноны (напр.,
убихиноны), каротиноиды, ретиноиды (рети-нол, ретиналь и др.), токоферолы, долихолы
(содержат 16-20 пренильных остатков, из к-рых концевой, несущий группу ОН, полностью
насыщен) и порфирины. Ок. 20% всей массы мембраны составляет прочно связанная
вода. С мембранами связываются также катионы, преим. Са2+ и Mg2+,
входящие в хелатные комплексы.
Важнейшая ф-ция М. б.-регуляция
обмена в-в между клеткой и средой, а также между разл. отсеками (компарт-ментами)
внутри самой клетки.
Липиды мембран. Осн.
липидные компоненты М.б.-фос-фолипиды, гликолипиды и стерины. Каждая группа
этих липидов представлена большим числом разнообразных соединений. Так, в мембране
эритроцитов человека содержится не менее 20 разл. представителей осн. фосфолипида
этой мембраны - фосфатидилхолина; в целом же в мембране эритроцитов идентифицировано
ок. 200 разл. липидов.
В клетках млекопитающих
плазматич. мембраны обогащены холестерином и гликосфииголипидами, тогда как
мембраны органоидов содержат эти липиды в малых ко-лвах. Наиб. распространенные
липиды, имеющие цвиттер-ионную структуру, в большинстве мембран клеток млекопитающих-фосфатидилхолин
и сфингомиелин (в митохондри-альных мембранах - фосфатидилэтаноламин). Дифосфати-дилглицерин
в значит. кол-вах присутствует только в мембранах митохондрий (в осн. в их внутр.
мембране). В плазматич.
мембранах содержание фосфатидилсерина обычно больше, чем фосфатидилинозита (фосфоинозитида),
для внутриклеточных мембран характерно обратное соотношение. В мембранах миелина
широко представлены цереб-розиды. Др. плазматич. мембраны содержат, как правило,
более сложные гликолипиды, такие, напр., как ганг-лиозиды. Фосфатидилэтаноламин
в мембранах миелина и тромбоцитов находится преим. в плазмалогеновой форме.
Мембраны клеток высших
растений и дрожжей по ли-пидному составу во многом сходны с соответствующими
мембранами клеток млекопитающих. Однако в них совсем нет сфингомиелина, а фосфатидилсерин
присутствует лишь в следовых кол-вах. Главные стерины мембран растит. клеток
- ситостерин и стигмастерин, мембран грибов и дрожжей - эргостерин и зимостерин.
Мембраны хлороплас-тов фотосинтезирующих растений и синезеленых водорослей близки
по своему липидному составу и содержат моно-и дигалактозилдиацилглицерины, 6-сульфохиновозилдиа-цилглицерин
и фосфатидилглицерин.
Мембраны бактерий, как
правило, имеют более простой липидный состав, чем мембраны растит. и животных
клеток. Все бактерии, за исключением микоплазм, не содержат стеринов. Фосфолипиды
мембран грамположит. бактерий представлены гл. обр. фосфатидилглицерином и его
ами-ноациальными производными, а также дифосфатидилгли-церином. В небольшом
кол-ве в этих мембранах нередко встречается фосфатидилинозит. У грамотрицат.
микроорганизмов в составе мембранных фосфолипидов преобладает Фосфатидилэтаноламин.
Фосфатидилхолин в бактериальных мембранах либо совсем не содержится, либо присутствует
в малых кол-вах. Содержание фосфатидилсерина в этих мембранах обычно также незначительно.
Широко представлены в бактериальных мембранах разл. гликозил-диацилглицерины.
Осн. компоненты мембран
оболочечных вирусов (вирус гриппа, лейковирусы, вирус стоматита), как и плазматич.
мембран клеток животных,-фосфатидилхолин, сфингомие-лин, Фосфатидилэтаноламин
и холестерин.
Липидный состав клеточных
мембран изменчив. В меньшей степени это проявляется в животных клетках, находящихся
в условиях стабильной внутр. среды. Однако и в этом случае можно модифицировать
состав липидов в нек-рых мембранах, меняя пищ. рацион. Липидный состав мембран
растений заметно изменяется в зависимости от освещенности, т-ры и рН. Еще более
изменчив состав бактериальных мембран. Он варьирует не только в зависимости
от штамма, но и в пределах одного и того же штамма, а также от условий культивирования
и фазы роста. У вирусов, имеющих липопротеиновую оболочку, липидный состав мембран
также не постоянен и определяется составом липидов клетки-хозяина.
Липиды-осн. строит. материал,
из к-рого формируются клеточные мембраны. Сложность, многообразие и изменчивость
липидного состава мембран позволяет предположить, что они участвуют также в
регуляции важнейших мембранных процессов.
Мембранные белки. Мол.
масса мембранных белков обычно варьирует в пределах от 10 тыс. до 240 тыс. Они
значительно различаются между собой по прочности связывания с мембраной. Белки,
наз. периферич. или поверхностными, сравнительно слабо связаны с мембраной и
отделяются от нее в мягких условиях, напр. в р-рах, имеющих высокую ионную силу
или содержащих комплексоны. Намного прочнее связаны с мембраной т. наз. интегральные,
или внутримембранные, белки (см. рис.). Чтобы их выделить, требуется, как правило,
предварительно разрушить мембрану с помощью ПАВ или орг. р-рителей.
Периферич. белки по своим
св-вам мало отличаются от обычных водорастворимых белков. Характерная особенность
интегральных белков - плохая р-римость в воде и склонность к образованию ассоциатов.
Их удается перевести в
р-р при добавлении ПАВ, иногда с помощью орг. р-рителей (напр., 2-хлорэтанола,
бутанола, ДМФА).
Особенность интегральных
белков - наличие в их поли-пептидной цепи довольно протяженных участков с преобладающим
содержанием неполярных аминокислот. Как правило, эти участки имеют конформацию
a-спирали, на наружной стороне к-рой расположены боковые углеводородные фрагменты
аминокислотных остатков, в результате чего вся спираль, в целом, приобретает
гидрофобный характер. Доля a-спиральных участков в мембранных белках довольно
велика (составляет 30-50%), остальная часть полипептид-ной цепи находится преим.
в форме неупорядоченного клубка. Участков с b-структурой, как правило, мало.
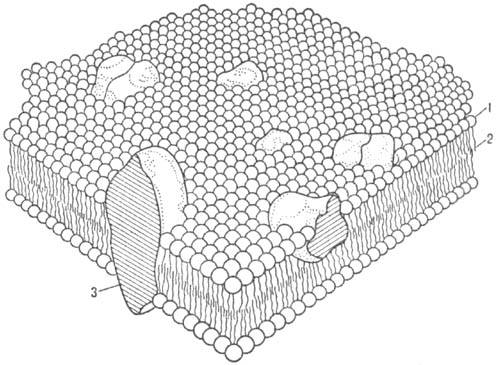
Схема мозаичной модели
клеточной мембраны: 1 -полярная
головка молекулы липида; 2 - углеводородная цепь
молекулы липида; 3 - интегральный белок.
Гидрофобные a-спиральные
участки интегральных белков обычно содержат от 17 до 26 аминокислотных остатков,
что вполне достаточно, чтобы полипептидная цепь однократно пересекла М. б. В
белках, к-рые пронизывают М. б. насквозь, такие гидрофобные тяжи соединяют между
собой полярные области белковой молекулы, находящиеся на противоположных сторонах
мембраны. У белков, расположенных только на одной стороне М.б. и погруженных
в нее лишь частично, a-спирали служат своеобразным гидрофобным "якорем",
прочно удерживающим белок в мембране. В нек-рых случаях "заякоривание"
белков в М.б. происходит при помощи ковалентно связанных с ними липидов.
Типичные примеры белков,
к-рые удерживаются в М.б. благодаря гидрофобному a-спиральному участку полипеп-тидной
цепи,-цитохром b5-редуктаза и цитохром b5.
К белкам, полипептидная цепь к-рых однократно пересекает М. б., относятся, напр.,
антигены тканевой совместимости и мембраносвязанные иммуноглобулины, к белкам,
пересекающим М. б. более одного раза,-бактериородопсин. Нередко мембранные белки
представляют собой сложные комплексы, состоящие из неск. субъединиц (напр.,
цитохром с-ок-сидаза состоит из 12 субъединиц).
Мембранные белки наряду с липидами играют важную структурную роль, кроме этого они ответственны за выполнение подавляющего большинства специализир. ф-ций отдельных мембран. Они служат катализаторами протекающих в мембранах и на их пов-сти р-ций (см., напр., Дыхание), участвуют в рецепции гормональных и антигенных сигналов и т.п. (см., напр., Аденилатциклаза), выполняют транспортные ф-ции, обеспечивают пиноцитоз (захват клеточной пов-стью и поглощение клеткой жидкости), хемотаксис (перемещение клетки, обусловленное градиентом концентраций к.-л. в-ва в среде) и т.п. Мн. из периферич. белков-компоненты цитоскелета (совокупность филаментов и микротрубочек цитоплазмы) и связанных с ним сократит. элементов, к-рые обусловливают форму клетки и ее движение.
Ферментативная активность
присуща мн. мембраносвя-занным белкам, причем мембраны разл. клеток и отдельных
органоидов имеют свой характерный набор ферментов. Как правило, ферментные белки
располагаются в М. б. в определенном порядке, к-рый делает возможным последовательное
протекание р-ций метаболии, цикла.
Молекулярная организация
мембран. Структурная основа М. б.-липидный бислой. В продольной плоскости
М.б. представляет собой сложную мозаику из разнообразных липидов и белков, причем
их распределение по пов-сти М.б. неоднородно. В нек-рых М. б. имеются обширные
участки липидно-го бислоя, практически свободные от белков (напр., в эритроцитах
белки занимают только 35% площади пов-сти всей М.б., в микросомах-23%). При
высоком содержании белка в М.б. липиды не образуют сплошной бислой, а располагаются
в виде отдельных вкраплений между белковыми молекулами. Сам липидный бислой
в мембране может иметь доменную структуру в результате, напр., сосуществования
несмешиваемых липидных фаз, находящихся в двух разл. физ. состояниях - гелевом
и жидкокристаллическом. Часть липидов в М. б. может находиться также в составе
т. наз. небислойных фаз (мицеллярная фаза, гексагон. фаза и др.). Ассоциации
липидов в М.б. способствует также их взаимод. с многозарядными катионами (Са2
+ , Mg2+ и др.), периферич. белками, нек-рыми мембраноактивными
в-вами (напр., гормонами).
Специфич. взаимод. между
отдельными белками приводят к тому, что в М. б. образуются белковые ассоциаты,
или ансамбли, к-рые по составу и св-вам отличаются от окружающих участков мембраны
и часто окружены липидами определенного типа. Иногда липопротеиновые участки
М.б., содержащие характерный набор белков и липидов, удается выделить при фрагментации
мембран. Образование ассоциатов белков может происходить также в результате
их специфич. связывания на пов-сти М. б. с нек-рыми водорастворимыми белками
(напр., с антителами, лектинами) или при фазовом переходе липидов в мембране
(обычно белки скапливаются там, где липиды продолжают оставаться в жидкокристаллич.
состоянии).
Неоднородность М. б. связана
также со структурными и функцион. различиями наружной и внутр. сторон мембраны,
обусловленными неодинаковым распределением отдельных компонентов (белков, липидов,
углеводов и др.). Характерный пример асимметрич. распределения липидов - плаз-матич.
мембрана эритроцитов. Холинсодержащие фосфоли-пиды (фосфатидилхолин и сфингомиелин)
преобладают у них на наружной стороне мембраны, а фосфатидилэтанол-амин, фосфатидилсерин
и фосфатидилинозит связаны пре-им. с ее внутр. пов-стью, обращенной в сторону
цитоплазмы. Сходное распределение фосфолипидов обнаружено в плазматич. мембранах
др. животных клеток.
Если асимметрия в расположении
липидов в большинстве случаев в М.б. носит относит. характер (т.е. на наружной
и внутр. стороне мембраны находятся обычно одни и те же липиды, хотя и в разной
концентрации), то асимметрия в расположении белков является абсолютной - все
молекулы данного белка определенным образом расположены в мембране. Так, цитохром
b5 всегда локализован только на цитоплазматич. стороне мембраны
эндоплазматич. ретику-лума. В случае проникающего через мембрану эритроцитов
белка гликофорина (ответствен за мн. ф-ции, в т. ч. препятствует слипанию эритроцитов)
N-конец полипептидной цепи, содержащий ковалентно связанные углеводы, находится
на наружной пов-сти, а С-конец-на цитоплазматич. стороне мембраны. Строго определенную
ориентацию в М. б. имеют все молекулы бактериородопсина, у к-рого полипептидная
цепь неск. раз пересекает липидный бислой, а также сложные белковые комплексы,
состоящие из неск. субъединиц (напр., цитохромоксидаза, аденилатциклаза).
Отдельные компоненты М.
б. могут менять свое взаимное расположение, перемещаться в ней на значит. расстояния,
а также покидать мембрану или внедряться в нее в ходе разл. метаболич. процессов.
Такая динамичность позволяет М.б. быстро
адаптироваться к изменению условий окружающей среды и оперативно откликаться
на разнообразные внеш. сигналы и стимулирующие воздействия.
Динамич. св-ва М.б. обусловлены
текучестью липидного бислоя, гидрофобная область к-рого в жидкокристаллич. состоянии
имеет микровязкость, сравнимую с вязкостью легкой фракции машинного масла. Поэтому
молекулы липидов, находящиеся в бислое, обладают довольно высокой подвижностью
и могут совершать разнообразные движения-поступательные, вращательные и колебательные.
В случае липидов большой
вклад в подвижность дают внутримол. движения углеводородных цепей. Они происходят
путем гош-транс-поворотов (см. Конформационный анализ)смежных
звеньев углеводородной цепи вокруг связи С—С. Благодаря высокой конформац. подвижности
цепей в них постоянно возникают изгибы и изломы, что приводит к нарушению регулярного
расположения липидных молекул в бислое и к появлению в нем дефектов упаковки,
называемых "кинки" и "джогги".
Внутримол. подвижность
разл. участков липидной молекулы, находящейся в бислое, неодинакова. Наим. подвижностью
обладает глицериновый остов молекулы, к-рый служит как бы жестким "якорем",
ограничивающим движения близлежащих участков углеводородных цепей. По направлению
к середине бислоя подвижность цепей возрастает и становится максимальной в области
концевых ме-тильных групп. Довольно высокой недвижностью обладает также полярная
головка липидной молекулы.
Помимо движений отдельных
участков липидной молекулы относительно друг друга в жидкокристаллич. бислое
происходят также движения всей молекулы как единого целого. Они включают: аксиальное
вращение молекулы вокруг ее длинной оси, перпендикулярной к плоскости бислоя,
маятниковые и поплавочные колебания молекулы относительно ее равновесного положения
в бислое, перемещение молекулы вдоль бислоя (латеральная диффузия) и перескок
ее с одной стороны бислоя на другой (флип-флоп). Все эти движения совершаются
с разными скоростями.
Аксиальное вращение липидных
молекул происходит очень быстро с частотой порядка 107-108с-1,
тогда как латеральная диффузия осуществляется гораздо медленнее. Тем не менее
при среднем коэф. латеральной диффузии липидов ок. 10-8см2.с-1,
измеренном для мн. М.б., липидной молекуле потребуется всего 1 с, чтобы промигрировать
от одного конца клетки до другого. Очень медленно протекает в липидном бислое
флип-флоп. Обычно полупериод флип-флопа составляет величины порядка неск. часов
или даже дней. Однако в нек-рых мембранах скорость флип-флопа м. б. значительно
выше (полупериод 1-2 мин), что объясняется участием определенных интегральных
белков в переносе липидных молекул через мембрану.
Иммобилизация липидов может
происходить в результате латерального фазового разделения, приводящего к образованию
гелевой фазы, или при их взаимод. с белками. Предполагается, что интегральные
белки окружены пограничным слоем липидных молекул (т. наз. аннулярные липиды),
подвижность к-рых ограничена или, по крайней мере, нарушена в результате контакта
с неровной пов-стью белковой глобулы.
Внутримол. динамика мембранных
белков изучена меньше, чем липидов. Известно лишь, что боковые заместители на
тех участках полипептидной цепи, к-рые погружены в липидный бислой, в значит.
мере иммобилизованы. Мн. мембранные белки способны легко диффундировать вдоль
мембраны и обладают довольно высокой вращат. подвижностью. Но даже в случае
самых подвижных белков измеряемые коэф. диффузии примерно на порядок ниже, чем
для липидных молекул. Времена вращат. релаксации для интегральных белков лежат
в диапазоне от 20 до 500 мкс, а коэф. латеральной диффузии (вдоль бислоя) варьирует
от 7.10-9 до 10-12см2.с-1.
Быстрая диффузия белков
вдоль мембраны наблюдается только в жидкокристаллич. бислое, в гелевой фазе
белки не мигрируют.
Мобильными являются 20-50% мембранных белков, остальные имеют ограниченную подвижность
или совсем неподвижны. Причиной иммобилизации интегральных белков в мембране
м. б. их ассоциация с образованием крупных агрегатов или даже двухмерных кристаллич.
структур, взаимод. с периферич. белками, связывание с элементами цитоскелета
и т.п.
Исследования М.б. представляют
собой важную, активно развивающуюся область совр. биологии. С успехами в области
изучения мембран связаны мн. достижения в медицине, напр. установление механизмов
возникновения не-к-рых сердечно-сосудистых заболеваний и поиск подходов к их
лечению. Идеи и методы, возникшие при исследовании мембран, находят широкое
применение в онкологии, технологии создания искусств. органов, в трансплантац.
иммунологии, эмбриологии и др. Знание процессов, происходящих в мембранах, играет
важную роль в развитии таких направлений, как биоэнергетика и поиск эффективных
путей утилизации солнечной энергии, создание биосенсорных устройств, мембранная
технология и др.
Лит.: Ивков В. Г., Берестовский Г. Н., Динамическая структура ли-пидного бислоя, М., 1981; Бергельсон Л. Д., Мембраны, молекулы, клетки, М., 1982; Ивков В. Г., Берестовский Г. Н., Липидный бислой биологических мембран, М., 1982; Кагава Я., Биомембраны, пер. с япон., М., 1985; Сим Э., Биохимия мембран, пер. с англ., М., 1985; Болдырев А. А., Введение в биохимию мембран, М., 1986; Биологические мембраны, под ред. Дж. Б. С. Финдлея и В. X. Эванза, пер. с англ., М., 1990. Л. И. Барсуков.