

Словарь научных терминов
Матричные рибонуклеиновые кислоты
МАТРИЧНЫЕ РИБОНУКЛЕИНОВЫЕ КИСЛОТЫ (информационные РНК, мРНК, иРНК), молекулы РНК, представляющие собой комплементарные (см. Комплементарность) копии участков значащих цепей генов (ДНК), в к-рых закодирована информация об аминокислотных последовательностях полипептидных цепей белков. Образуются в результате транскрипции (синтез РНК на ДНК-матрице с помощью фермента ДНК-зависимой РНК-полимеразы) и послед. процессинга первичного транскрипта - удаления избыточных РНК-последовательностей с обоих концов молекулы и некодирующих последовательностей РНК, а также сшивания кодирующих участков РНК друг с другом. мРНК переносят генетич. информацию от генов к рибосомам, где происходит трансляция (синтез полипептидных
цепей на мРНК-матрице). При этом одну цепь мРНК могут одновременно транслировать неск. рибосом (т. наз. полирибосомы, или полисомы).
мРНК всегда состоят из одной полинуклеотидной цепи, т. е. относятся к однотяжевым РНК. Как и любые др. РНК, мРНК включают остатки адениловой (А), гуаниловой (G), уридиловой (U) и цитидиловой (С) к-т, связанных между собой 3'-5'-фосфодиэфирными связями (ф-ла I). Вторичная структура мРНК представлена многочисл. двухспиральными участками ("шпильками"), образующимися в результате комплементарного спаривания оснований (А с U и G с С) одной и той же цепи; шпильки соединены между собой короткими однотяжевыми участками. При связывании с рибосомой макромолекулярная структура мРНК существенно изменяется, а двухспиральные ее участки в районе непосредств. контакта с рибосомой переходят в однотяжевое состояние.
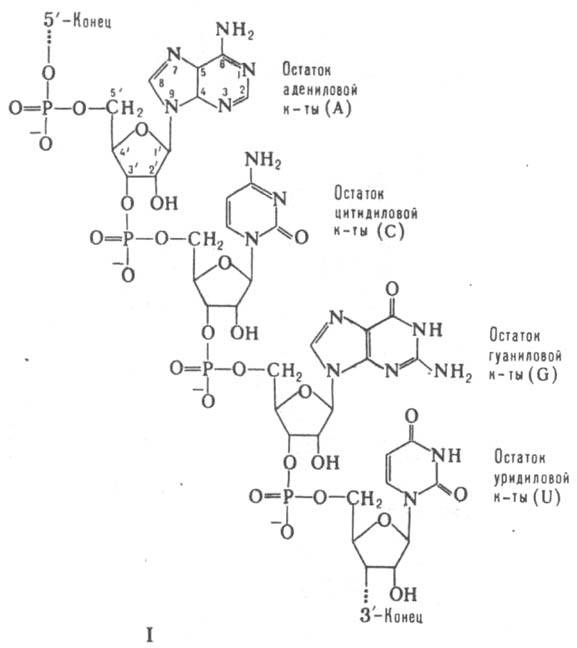
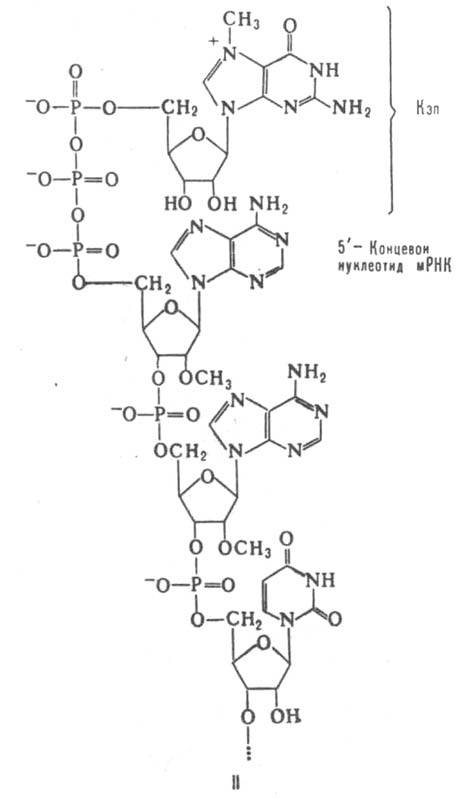
мРНК составляют незначит. долю (3-5%) от суммарной клеточной РНК. В клетке бактерий мРНК весьма нестабильны и быстро разрушаются: время их полужизни 2-3 мин. В отличие от них, мРНК в клетках эукариот (все организмы, за исключением бактерий и синезеленых водорослей), как правило, сохраняются в течение неск. часов. В значит. мере это связано с тем, что в эукариотич. клетках мРНК всегда находятся в комплексе с белками, образуя матричные рибонуклеопротеидные частицы, или информосомы. Белковый состав информосом в период, предшествующий трансляции, и в момент трансляции (т.е. в составе полисом) различен. Молекулы мРНК содержат два типа нуклеотидных последовательностей - транслируемые и нетранслируемые. Транслируемый участок мРНК представляет собой совокупность тринуклеотидов (кодонов; см. Генетический код), кодирующих полипептидную цепь. Он начинается с инициирующего кодона (в подавляющем числе случаев AUG) и заканчивается терминирующим (UAG, UAA или UGA). Одна молекула мРНК прокариот (бактерии и синезеленые водоросли) часто кодирует две или более разных полипептидных цепи (такие мРНК наз. полицистронными) и содержит соотв. неск. кодирующих последовательностей. Аналогичным образом организованы РНК многих РНК-содержащих бактериофагов (вирусы бактерий), к-рые в клетках бактерий выполняют ф-ции мРНК. В полицистронных мРНК последовательности, кодирующие полипептидные цепи, разделены межцистронными нетранслируемыми участками (спейсерами), длина к-рых варьирует в широких пределах. Между инициирующим кодоном ближайшей к 5'-концу мРНК транслируемой последовательности и этим концом обычно находится 50-200 нуклеотидных остатков. 3'-Конец мРНК отделен от терминирующего кодона ближайшей к нему кодирующей последовательности, как правило, значительно более длинным нетранслируемым участком (до неск. тыс. нуклеотидных остатков). Эти нетранслируемые области, по-видимому, необходимы для создания специфич. макромолекулярной структуры мРНК, а также содержат т. наз. сигналы инициации трансляции. наиб. хорошо изучена такая сигнальная последовательность у мРНК прокариот (открыта Дж. Шайн и Л. Дальгарно и носит их имя), содержащая 3-7 нуклеотидных остатков, расположенных перед инициирующим ко-доном (т. е. ближе к 5'-концу молекулы) каждой транслируемой последовательности. Последовательность Шайн-Дальгарно комплементарна 3'-концевому участку РНК малой субчастицы рибосом и, по всей вероятности, образует с ним в процессе инициации трансляции двухспиральный комплекс. Вторичная структура мРНК в районе сигнальной последовательности и инициирующего кодона оказывает влияние на эффективность инициации трансляции. Наличие или отсутствие своб. 5'-концевого участка в мРНК прокариот несущественно для начала трансляции. В отдельных случаях рибосома способна начинать трансляцию внутр. кодирующих последовательностей. Нек-рые мРНК прокариот (чаще всего кодирующие ферменты биосинтеза аминокислот) кроме основных транслируемых последовательностей ближе к их 5'-концу содержат регуляторную последовательность, в к-рой закодирован короткий полипептид (этот район мРНК носит назв. аттенюатора). Синтез этого полипептида происходит во время образования мРНК и может приводить к преждевременному окончанию синтеза последней. У нек-рых РНК-содержащих вирусов эукариот в РНК закодировано большое число белков, которые в процессе трансляции синтезируются в виде гигантской единой полипептидной цепи (полипротеина), расчленяемой затем специфич. протеазами на отдельные белковые молекулы. У эукариот мРНК, как правило, моноцистронны. 5'-Конец таких мРНК часто несет модифицир. группировку (т. наз. кэп от англ. cap - головка, ф-ла II), присоединяемую к мРНК после завершения транскрипции с помощью спец. ферментной системы. 5'-Концевые остатки самой полинуклеотидной цепи мРНК, связанные с кэпом, также модифицированы -обычно они содержат остатки рибозы, метилированные по положению 2'. Наличие кэпа в мРНК повышает эффективность ее трансляции. Предполагают, что у эукариот рибосома первоначально связывается с 5'-концом молекулы, а затем "скользит" по цепи мРНК вплоть до инициирующего кодона транслируемой последовательности. Б. ч. молекул мРНК в эукариотич. клетке содержит на своем 3'-конце гомополимерную цепь (полиаденилат), в к-рой находится от 20 до 200 остатков адениловой к-ты. Полиаденилирование осуществляется ферментом полиаденилатполимеразой после окончания транскрипции как последний этап процессинга 3'-концевой области мРНК. Функцион. роль полиаденилатной последовательности в мРНК достоверно не установлена; возможно, она служит для связывания спец. белков в информосоме. Нек-рые мРНК (напр., гистоновые мРНК) полиаденилированы лишь частично. Благодаря наличию полиаденилатной последовательности мРНК м. б. легко отделены от остальных клеточных РНК путем избират. комплементарного связывания с иммобилизованными олигомерами уридиловой или тимидиловой к-ты.
Полиаденилатные последовательности на 3'-концах мРНК часто используют также для получения их ДНК-копий с помощью олигомера тимидиловой к-ты в качестве затравки и фермента обратной транскриптазы. Эти копии в свою очередь используют для получения индивидуальных генов методами генетич. инженерии. Существование мРНК предсказано в 1956-58 А. С. Спириным и А. Н. Белозерским, а также независимо от них Э. Волкиным и Ф. Астрачаном. Экспериментально мРНК открыты в 1961 в лабораториях М. С. Мезелсона, Дж. Д. Уотсона и С. Спигелмена. Лит.: Информационные макромолекулы, пер. с англ., М., 1965; Спирин А. С., Молекулярная биология. Структура рибосомы и биосинтез белка, М., 1986; Льюин Б., Гены, пер. с англ., М., 1987. А. А. Богданов.
мРНК составляют незначит. долю (3-5%) от суммарной клеточной РНК. В клетке бактерий мРНК весьма нестабильны и быстро разрушаются: время их полужизни 2-3 мин. В отличие от них, мРНК в клетках эукариот (все организмы, за исключением бактерий и синезеленых водорослей), как правило, сохраняются в течение неск. часов. В значит. мере это связано с тем, что в эукариотич. клетках мРНК всегда находятся в комплексе с белками, образуя матричные рибонуклеопротеидные частицы, или информосомы. Белковый состав информосом в период, предшествующий трансляции, и в момент трансляции (т.е. в составе полисом) различен. Молекулы мРНК содержат два типа нуклеотидных последовательностей - транслируемые и нетранслируемые. Транслируемый участок мРНК представляет собой совокупность тринуклеотидов (кодонов; см. Генетический код), кодирующих полипептидную цепь. Он начинается с инициирующего кодона (в подавляющем числе случаев AUG) и заканчивается терминирующим (UAG, UAA или UGA). Одна молекула мРНК прокариот (бактерии и синезеленые водоросли) часто кодирует две или более разных полипептидных цепи (такие мРНК наз. полицистронными) и содержит соотв. неск. кодирующих последовательностей. Аналогичным образом организованы РНК многих РНК-содержащих бактериофагов (вирусы бактерий), к-рые в клетках бактерий выполняют ф-ции мРНК. В полицистронных мРНК последовательности, кодирующие полипептидные цепи, разделены межцистронными нетранслируемыми участками (спейсерами), длина к-рых варьирует в широких пределах. Между инициирующим кодоном ближайшей к 5'-концу мРНК транслируемой последовательности и этим концом обычно находится 50-200 нуклеотидных остатков. 3'-Конец мРНК отделен от терминирующего кодона ближайшей к нему кодирующей последовательности, как правило, значительно более длинным нетранслируемым участком (до неск. тыс. нуклеотидных остатков). Эти нетранслируемые области, по-видимому, необходимы для создания специфич. макромолекулярной структуры мРНК, а также содержат т. наз. сигналы инициации трансляции. наиб. хорошо изучена такая сигнальная последовательность у мРНК прокариот (открыта Дж. Шайн и Л. Дальгарно и носит их имя), содержащая 3-7 нуклеотидных остатков, расположенных перед инициирующим ко-доном (т. е. ближе к 5'-концу молекулы) каждой транслируемой последовательности. Последовательность Шайн-Дальгарно комплементарна 3'-концевому участку РНК малой субчастицы рибосом и, по всей вероятности, образует с ним в процессе инициации трансляции двухспиральный комплекс. Вторичная структура мРНК в районе сигнальной последовательности и инициирующего кодона оказывает влияние на эффективность инициации трансляции. Наличие или отсутствие своб. 5'-концевого участка в мРНК прокариот несущественно для начала трансляции. В отдельных случаях рибосома способна начинать трансляцию внутр. кодирующих последовательностей. Нек-рые мРНК прокариот (чаще всего кодирующие ферменты биосинтеза аминокислот) кроме основных транслируемых последовательностей ближе к их 5'-концу содержат регуляторную последовательность, в к-рой закодирован короткий полипептид (этот район мРНК носит назв. аттенюатора). Синтез этого полипептида происходит во время образования мРНК и может приводить к преждевременному окончанию синтеза последней. У нек-рых РНК-содержащих вирусов эукариот в РНК закодировано большое число белков, которые в процессе трансляции синтезируются в виде гигантской единой полипептидной цепи (полипротеина), расчленяемой затем специфич. протеазами на отдельные белковые молекулы. У эукариот мРНК, как правило, моноцистронны. 5'-Конец таких мРНК часто несет модифицир. группировку (т. наз. кэп от англ. cap - головка, ф-ла II), присоединяемую к мРНК после завершения транскрипции с помощью спец. ферментной системы. 5'-Концевые остатки самой полинуклеотидной цепи мРНК, связанные с кэпом, также модифицированы -обычно они содержат остатки рибозы, метилированные по положению 2'. Наличие кэпа в мРНК повышает эффективность ее трансляции. Предполагают, что у эукариот рибосома первоначально связывается с 5'-концом молекулы, а затем "скользит" по цепи мРНК вплоть до инициирующего кодона транслируемой последовательности. Б. ч. молекул мРНК в эукариотич. клетке содержит на своем 3'-конце гомополимерную цепь (полиаденилат), в к-рой находится от 20 до 200 остатков адениловой к-ты. Полиаденилирование осуществляется ферментом полиаденилатполимеразой после окончания транскрипции как последний этап процессинга 3'-концевой области мРНК. Функцион. роль полиаденилатной последовательности в мРНК достоверно не установлена; возможно, она служит для связывания спец. белков в информосоме. Нек-рые мРНК (напр., гистоновые мРНК) полиаденилированы лишь частично. Благодаря наличию полиаденилатной последовательности мРНК м. б. легко отделены от остальных клеточных РНК путем избират. комплементарного связывания с иммобилизованными олигомерами уридиловой или тимидиловой к-ты.
Полиаденилатные последовательности на 3'-концах мРНК часто используют также для получения их ДНК-копий с помощью олигомера тимидиловой к-ты в качестве затравки и фермента обратной транскриптазы. Эти копии в свою очередь используют для получения индивидуальных генов методами генетич. инженерии. Существование мРНК предсказано в 1956-58 А. С. Спириным и А. Н. Белозерским, а также независимо от них Э. Волкиным и Ф. Астрачаном. Экспериментально мРНК открыты в 1961 в лабораториях М. С. Мезелсона, Дж. Д. Уотсона и С. Спигелмена. Лит.: Информационные макромолекулы, пер. с англ., М., 1965; Спирин А. С., Молекулярная биология. Структура рибосомы и биосинтез белка, М., 1986; Льюин Б., Гены, пер. с англ., М., 1987. А. А. Богданов.
-метилацетофенон
-метоксиацетофенон
2-меркаптобензотиазол
2-меркаптоэтиламин
2-метил-5-винилпиридин
N-метилпирролидон
Магнезоны
Магнетохимия
Магниевые удобрения
Магний
Магнийорганические соединения
Магнитная восприимчивость
Магнитная постоянная
Магнитно-спиновые эффекты
Магнитные материалы
Магнитный момент
Магния галогениды
Магния гидроксид
Магния карбонат
Магния нитрат
Магния оксид
Магния перхлорат
Магния сплавы
Магния сульфат
Мазут
Майзенхаймера перегруппировка
Мак-лафферти перегруппировка
Мак-фадьена-стивенса реакция
Макарова фазы
Маковое масло
Макро- и микрокомпоненты
Макрокинетика
Макролиды
Макромолекула
Макромономеры
Макропористые ионообменные смолы
Макрорадикалы
Максимальная работа реакции
Малапрада реакция
Малахитовый зеленый
Малеиновая и фумаровая кислоты
Малеиновый ангидрид
Малоновая кислота
Малоновый эфир
Малононитрил
Мальтены
Мальтоза
Мальтол
Манганаты
Манганин
Маннаны
Маннит
Манниха реакция
Манноза
Маноилоксиды
Манометры
Маноол
Марганец
Марганецорганические соединения
Марганца карбонат
Марганца карбонилы
Марганца нитрат
Марганца оксиды
Марганца сульфат
Марганцевые удобрения
Маскирование
Маслонаполненные каучуки
Маслостойкость
Масляная кислота
Масляные краски
Масляные лаки
Масляный альдегид
Масс-спектрометрия
Массовое число
Массообмен
Мастики
Мастикс
Масштабный переход
Маточные средства
Матрица плотности
Матричные рибонуклеиновые кислоты
Матричный синтез
Машинные масла
Меди ацетаты
Меди гидроксиды
Меди карбонаты
Меди нитрат
Меди оксиды
Меди сплавы
Меди сульфат
Меди сульфиды
Меди хлориды
Медицинские масла
Медноаммиачные волокна
Медные удобрения
Медь
Медьорганические соединения
Меервейна реакция
Межгалогенные соединения
Межкристаллитная коррозия
Межмолекулярные взаимодействия
Межфазная поликонденсация
Межфазные скачки потенциала
Межфазный катализ
Мезаконовая кислота
Мезидин
Мезитила окись
Мезитилен
Мезо
Мезоионные соединения
Мезоксалевая кислота
Мезомерия
Мезомерный эффект
Мезонная химия
Мейера - шустера перегруппировка
Мейера реакция
Меламин
Меланины
Мелем
Мельхиор
Мембранный катализ
Мембранный потенциал
Мембраны биологические
Мембраны жидкие
Мембраны ионообменные
Мембраны разделительные
Менделевий
Ментадиены
Ментаны
Ментены
Ментол
Ментон
Меншуткина реакция
Мепробamat
Меркаптаны
Меркаптохинолины
Меркуриметрия
Мерсеризация
Мета
Метаболизм
Метакриламид
Метакриловая кислота
Метакрилонитрил
Металепсия
Металлиды
Металлизация полимеров
Металлилхлорид
Металлирование
Металлическая связь
Металлические волокна
Металлические кристаллы
Металлические радиусы
Металлические соединения
Металлов окисление
Металлокомплексный катализ
Металлопласты
Металлополимеры
Металлопротеиды
Металлотермия
Металлоцены
Металлургия
Металлы
Металлы органические
Метальдегид
Метан
Метанол
Метансульфокислота
Метансульфохлорид
Метатезис
Метафосфаты органические
Метил-b-нафтилкетоh
Метилакрилат
Метилаль
Метиламины
Метилацетат
Метилацетилен
Метилбензолсульфонат
Метилвинилкетон
Метилдихлорфосфат
Метилдихлорфосфин
Метилдихлорфосфит
Метилдихлорфосфонат
Метилдофа
Метиленовый голубой
Метиленхлорид
Метилиафталины
Метилизобутилкетон
Метилизотиоцианат
Метилизоцианат
Метилметакрилат
Метилнонилацетальдегид
Метиловый спирт
Метилсерная кислота
Метилстиролы
Метилтетрафторфосфоран
Метилтимоловый синий
Метилфторид
Метилхлорид
Метилхлорсиланы
Метилцеллюлоза
Метилэтилбензолы
Метилэтилкетон
Метиновые красители
Метионин
Метионинметилсульфонийхлорид
Механизм реакции
Механические процессы
Механические свойства
Механохимия
Меченые атомы
Меченые соединения
Мешалки
Микотоксины
Микробиологический синтез
Микроволновая спектроскопия
Микрография
Микрокапсулирование
Микрокристаллоскопия
Микроудобрения
Микрофильтрация
Микрохимический анализ
Микроэлементы
Микроэмульсии
Миллона реакция
Минерал
Минерализация
Минеральные воды
Минеральные удобрения
Минорные нуклеозиды
Миоглобин
Миозин
Мирцен
Мирценаль
Митомицины
Михаэлиса-беккера реакция
Михаэля реакция
Михлера кетон
Мицеллирный катализ
Мицеллообразование
Мицеллы
Мицеллярные системы
Мицунобу реакция
Многокомпонентные системы
Многофотонные процессы
Мовеин
Модакриловые волокна
Моделирование
Модификация белков
Модифицирование древесины
Модифицирование полимеров
Молекула
Молекулярная биология
Молекулярная динамика
Молекулярная масса
Молекулярная масса полимера
Молекулярная механика
Молекулярность реакции
Молекулярные интегралы
Молекулярные комплексы
Молекулярные кристаллы
Молекулярные модели
Молекулярные соединения
Молекулярные спектры
Молекулярный анализ
Молибдаты
Молибден
Молибдена карбонилы
Молибдена оксиды
Молибдена сплавы
Молибдена фториды
Молибдена хлориды
Молибденовые удобрения
Моллюскоциды
Молочная кислота
Моляльность
Молярность
Монель-металл
Моноаминоксидазы
Моноглим
Монокристаллов выращивание
Монокристаллы
Мономеры
Мономолекулярные реакции
Мономолекулярный слой
Мононить
Моносахариды
Монофенолмонооксигеназы
Монохлорукссусная кислота
Моноэтаноламин
Морин
Морозостойкость
Морская коррозия
Морфин
Морфинановые алкалоиды
Морфолин
Морфотропия
Моторные масла
Моторные топлива
Мочевина
Мочевины цикл
Мощность дозы
Моющее действие
Мукайямы реакция
Мукополисахариды
Мультиплетность
Мумия
Муравьиная кислота
Муравьиный альдегид
Мурексид
Мускусы
Мутагены
Мутаротация
Мутации
Мыла
Мылонафт
Мышьяк
Мышьяка гидрид
Мышьяка хлориды
Мышьякорганические соединения
Мюон
Мюоний
Мягчители
Мёссбауэровская спектроскопия